


| name | Amanita suballiacea |
| name status | nomen acceptum |
| author | (Murrill) Murrill |
| english name | "Garlic-Odored Death Cap" |
| images |
 1. Amanita suballiaceae, South Fork Nature Ctr., Van Buren Co., Arkansas, U.S.A. |
| intro | The following description is based on the minimal original description and on original research by RET. This is one of the species that will often turn bright yellow in response to a drop of a strong base (like 5-10%KOH) on the cap. |
| cap | According to Murrill's (1941) original description, the cap of Amanita suballiacea is 30 - 40 mm wide, plano-convex, smooth, white, slightly viscid, with a nonstriate margin. The flesh is thin, white, and unchanging. |
| gills | The gills are "just" free and not connected to the stem by a line, crowded, white, unchanging. |
| stem | The stem is up to 90 × 5 - 10 mm, slightly expanded at the top, solid, white, with a thin membranous patch of limbate volva at the top of the bulb. The ring on the stem is white, membranous, thin, weak, apical or nearly so, and "hanging like a wet skirt." The stem's bulb is ellipsoid and may be deeply buried in the substrate. |
| odor/taste | The species is reported to have a strong odor of garlic which persists for some time in dried material. As a precautionary measure, this species should be considered deadly POISONOUS. |
| spores | The spores measure (7.2-) 7.5 - 9.5 (-9.8) × 6.5 - 8.0 (-8.2) µm and are subglobose to broadly ellipsoid to barely ellipsoid (infrequently globose) and weakly amyloid. Clamps are absent at base of basidia. [Note: The spore dimensions provided by Jenkins (1986) are apparently a typographical error. Curiously, Murrill also misdescribed the spores in his original description—as globose and 6 µm in diameter. The dimensions given above are from my study of the type.] |
| discussion |
The species was originally described from Florida,
USA and is reported to have been associated with liveoak and loblolly
pine. The species may occur in the states along the US coast
of the Gulf of Mexico as well as in states overlapping the Mississippi
River valley. Among other amanitas of the Atlantic Coastal Plain of North America, the odor of garlic is also present in A. alliacea (Murrill) Murrill—a white species with a small limbate volva presently assigned to Amanita sect. Lepidella. Among the collection of names of white taxa presently assigned to Amanita sect. Phalloideae in the eastern USA, the size and shape of the spores of A. suballiacea set it apart. Its spores are proportionately narrower than the spores of A. bisporigera G. F. Atk. and proportionately broader than those of A. magnivelaris Peck and the taxa of the A. elliptosperma G. F. Atk. "group." Whether other taxa with spores of this form exist is an open question, but no other name exists for such a species. The reader may want to examine the recently revised key to the taxa of sect. Phalloideae in North America.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita suballiacea | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| author | (Murrill) Murrill. 1941. Mycologia 33: 448. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| english name | "Garlic-Odored Death Cap" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| synonyms |
≡Venenarius suballiaceus Murrill. 1941. Mycologia 33: 437. The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MycoBank nos. | 284076, 291955 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| holotypes | FLAS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| type studies | Jenkins. 1979. Mycotaxon 10: 191. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of another researcher is based on uncompleted research by R. E. Tulloss. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileus | 30 - 40 mm wide, convex to planoconvex, slightly viscid, smooth, white, sordid at margin; context thin, white, unchanging on exposure; margin nonstriate, nonappendiculate; universal veil absent??. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamellae | just free, without decurrent lines on upper stipe, close, white, unchanging on bruising or with age, entire; lamellulae not described. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| stipe | 90 × 5 - 10 mm, white, subcylindric, smooth to subglabrous, slender; context not described; bulb [not present in holotype collection—ed.] ellipsoid, deeply buried; partial veil white, membranous, thin, weak, apical or just subapical, hanging like wet skirt; universal veil limbate, lobed, white(??), rather thick relative to stature of basidiocarp. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| odor/taste | Odor (holotype) strongly of garlic or onion when collected. Taste not recorded. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| macrochemical tests |
none recorded. 10% KOH - brilliant yellow on pileipellis and pileus context. Test vouchers: 21.ix.2014 Bunyard s.n., Justice AR-AM-29, and Nelson 10-31-82-EN1. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileipellis | from partial type study of RET: 40 - 50 µm thick, orange-brown in contrast to hyaline context, extensively gelatinized at surface; filamentous, undifferentiated hyphae 2.0 - 8.0 µm wide, subradially arranged in part, but with regions lacking dominant orientation, tightly interwoven in cross-section, branching; vascular hyphae not observed at surface. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| pileus context | from partial type study of RET: filamentous, undifferentiated hyphae 1.8 - 9.2 µm wide; acrophysalides dominating, clavate to broadly clavate to ??, up to ?? × ?? µm, thin-walled; vascular hyphae ?? µm wide. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamella trama | from partial type study of RET: not rehydratable in holotype | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| subhymenium | from partial type study of RET: not rehydratable in holotype | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| basidia | composite data from non-type material revised by RET: ?? × ?? μm, dominantly 4-sterigmate, infrequently 2-sterigmate, with sterigmata up to 8.0 μm long and having proportionately broad base (2.0± μm wide). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| universal veil | composite data from non-type material revised by RET: From limb on stipe's bulb, exterior layer: interwoven, gelatinized hyphae. From limb on stipe's bulb, interior: filamentous, undifferentiated hyphae 2.8 - 6.8 μm wide, plentiful, branching, containing minute particles; inflated cells plentiful, broadly clavate to ovoid, 60 - 76 × 35 - 50 μm, thin-walled, occasionally appearing to contain guttules; vascular hyphae 4.0 - 8.5 μm wide, branching. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| stipe context |
from partial type study of RET: longitudinally acrophysalidic; filamentous, undifferentiated hyphae 1.4 - 18.2 µm wide, with even those of largest diameter not having noticeably inflated segments; acrophysalides thin-walled, up to 201 × 62 µm; vascular hyphae 4.9 - 7.7 µm wide, uncommon. composite data from non-type material revised by RET: longitudinally acrophysalidic; filamentous, undifferentiated hyphae 2.5 - 9.5 μm wide, thin-walled; acrophysalides 80 - 209 × 34 - 36 μn, thin-walled; vascular hyphae up to 8.8± μm. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| lamella edge tissue | sterile. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
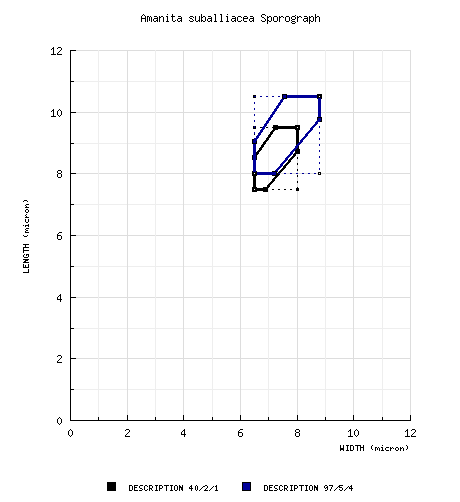
| basidiospores |
from type study of RET:
[40/2/1] (7.2-) 7.5 - 9.5
(-9.8) × 6.5 - 8.0 (-8.2) µm, (L = 8.7 -
8.9 µm; L’ = 8.8 µm; W = 7.2 - 7.3
µm; W’ = 7.2 µm; Q = (1.04-) 1.09 - 1.31
(-1.36); Q = 1.20 - 1.23; Q’ = 1.21),
hyaline, colorless, thin-walled, smooth, amyloid,
subglobose to broadly ellipsoid to occasionally
ellipsoid, somewhat adaxially flattened; apiculus
sublateral, small, cylindrical; contents
guttulate; ?probably whitish in
deposit. composite of data from all materials revised by RET: [97/5/4] (7.2-) 8.0 - 10.5 (-11.5) × 6.5 - 8.8 (-9.2) μm, (L = 8.7 - 10.1 μm; L' = 9.1 μm; W = 7.2 - 8.0 μm; W' = 7.5 μm; Q = (1.04-) 1.11 - 1.39 (-1.50); Q = 1.20 - 1.28; Q' = 1.22), colorless, hyaline, smooth, thin-walled, amyloid, subglobose to broadly ellipsoid to ellipsoid, rarely globose; apiculus sublateral, cylindric, proportionately small; contents guttulate; color in deposit not recorded. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ecology | Solitary or (frequently) in small groups. Arkansas: In mixed woods. Florida: In mixed woods including Pinus taeda and Quercus virginiana or under Serenoa repens with nearby Quercus. Pennsylvania: By Pinus. Texas: In sandy soil. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| material examined |
from protolog:
U.S.A.:
FLORIDA— Alachua Co. - Gainesville,
22.vi.1938 W. A. Murrill F 16495 (holotype,
FLAS). RET: U.S.A.: ARIZONA—Cochise Co. - 2.4 km northwest of Southwest Research Station, Onion Saddle Rd., Chiricahua Mycoflora Project site 33, 23.viii.1994 R. E. Tulloss 8-23-94-E [CMP 3403] (RET 678-1, nrLSU seq'd.; 4.2 km NW of Southwestern Research Station, road to Onion Saddle, Chiricahua Mycoflora Proj. site 37, 24.viii.1994 R. E. Tulloss 8-24-94-E [CMP 3961] (RET 676-2, nrLSU seq'd.). ARKANSAS—Montgomery Co. - Mt.? Ida, 31.x.1982 Edith Nelson s.n. [Tulloss 10-31-82-EN1] (RET 238-7). Van Buren Co. - South Fork Nature Center [35°33.697’ N/ 92°22.703’ W], 17.x.2010 Jay Justice AR-AM-29 (RET 458-10). CONNECTICUT—Middlesex Co. - East Haddam, Devil's Hopyard St. Pk., 4.ix.2011 Kevin Scalzo s.n. [Tulloss 9-4-11-K] (RET 490-1, nrITS seq'd.). FLORIDA—Alachua Co. - Gainesville, 22.vi.1938 W. A. Murrill F 16495 (holotype, FLAS). Charlotte Co. - Unkn. loc., 14.v.1997 Bruce Kuechmann s.n. (RET 260-1, nrLSU & mtLSU seq'd.). Lee Co. - Fort Myers, 3.v.2013 Benjamin Dion s.n. [mushroomobserver #132960] (RET 539-2, nrITS & nrLSU seq'd.). MAINE—Cumberland Co. - Standish, Rte. 114 & Plummer Rd. 0.4 km from Sebago Lk. [43.7528° N/ 70.5099° W, 93 m], 26.ix.1996 Samuel S. Ristich s.n. [Tulloss 9-26-96-SSR1] (RET 686-8, nrITS seq'd.). MICHIGAN—McComb Co. - Shelby Twp., ca. Washington, Stony Brook Metropolitan Pk. [42.73° N/ 83.07° W, 250-260 m], 30-31.viii.2011 Sandy Sheine s.n. (RET 491-7, nrITS seq'd.). NEW JERSEY—Monmouth Co. - Roosevelt, public school yard [40.2179° N/ 74.4748° W, 42 m], 26.viii.1993 R. E. Tulloss 8-26-93-A (RET 096-6, nrITS fragment seq'd.). OHIO—Unkn Co. - on border of Delaware & Franklin Cos., Westerville [40.1209º N/ 82.9262º W, 268 m], 10.x.2016 Michael McKnight s.n. (RET 744-3, nrITS-LSU seq'd.). PENNSYLVANIA—Butler Co. - Slippery Rock, Armco Pk. [41.0306° N/ 80.0216° W, 326 m], 21.viii.2014 Walt Sturgeon s.n. [mushroomobserver #174935] (RET 637-8, nrITS & nrLSU seq'd.). Clarion Co. - Clarion, St. Game Lands 63, Robert's Hill Rd. [41.1421° N/ 79.2830° W, 419 m], 6.viii.2011 Bill Bynum s.n. [Tulloss 8-6-11-C] (HKAS; RET 478-6, nrITS & nrLSU seq'd.). Luzerne Co. - Moon Lake Park, 6.x.2012 David Wasilewski s.n. [mushroomobserver #112828] (RET 524-8, nrITS & nrLSU seq'd.). TEXAS—Aransas Co. - Aransas Wildlife Refuge, 30.i.1989 Dr. Robert Harvey s.n. [Tulloss 1-30-89-AWR] (RET 244-1). WISCONSIN—Bayfield Co. - Cable [46.2019° N/ 91.2656° W, 400 m], 23.ix.2013 Britt Bunyard s.n. [mushroomobserver #146238] (RET 572-4, nrLSU seq'd.). ] Sauk Co. - Devil's Lake St. Pk. [43.4218° N/ 89.6972° W, 447 m], 28.ix.2013 B. Bunyard s.n. [mushroom observer #146910] (RET 631-5, nrITS & nrLSU seq'd.), 21.ix.2014 Britt Bunyard s.n. [mushroom observer #179727] (RET 671-6, nrITS seq'd.). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| discussion |
A note in Murrill's hand in the packet containing the
holotype indicates that the bases of the stems of the
two specimens comprising the type were lost. The type study of Jenkins (1979) is erroneous with regard to spore size and shape. It looks as though spore data from some other taxon were inadvertently substituted for the data from the present species. It is possible that Jenkins’ spore data for the present species somehow became swapped with his spore data for A. pseudoverna in (Jenkins 1979). In some correspondence and regional keys, RET once referred to this species as Amanita sp. T27. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| citations | —R. E. Tulloss | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.